单抗的定义:由一个B细胞克隆或其杂交瘤或通过分子生物学手段克隆构建单个抗体基因转染的细胞系产生的只能识别一种表位的高度均质性抗体。(此定义参考文献以及百度百科,若有异议请在本站论坛提出。)
1975年,Georges J. F. Kölhler(德国)和 César Milstein(英国和阿根廷国籍)发表了一篇文章:Continuous cultures of fused cells secreting antibody of predefined specificity(本站论坛可找到全文),这篇文章第一次阐述了单克隆抗体的制备(抗绵羊红细胞单克隆抗体),30多年过去了,这两位伟大的科学家发明的单抗制备方法在全世界范围内仍然广泛地应用着,两人于1984年与另一位科学家获得诺贝尔生理学和医学奖,他们的贡献是如此伟大,本人觉得就算给他们发三次诺奖也不为过,遗憾的是Georges J. F. Kölhler比较不幸,48岁时死于实验室火灾事故,这里也提醒各位读者注意实验室安全!
八卦完历史,接着说单抗。同多抗相比,单抗有如下优势:特异性好、重复性高、均一性强、应用更加广泛,但是与此同时,单抗制备流程长,首次制备成本高(重复制备成本低)、对技术要求也更高。根据前面的定义,制备单抗精华在于需要制备出一个能够稳定无限遗传的能分泌抗体的B细胞。于是不难理解单抗的两个基本问题:需要一个能分泌抗体的B细胞,能够稳定遗传。前一个问题,事实上就是免疫动物的问题,后一个问题,就是将B细胞与瘤细胞融合筛选的问题。免疫动物的问题和制备多抗大同小异,后一个问题才是制备单抗的核心问题。
当成功免疫动物后,便可杀死动物取得脾脏,里面含有我们需要的能分泌抗体的B细胞(不能无限增殖),再与瘤细胞融合,这样得到的杂交瘤既可以分泌抗体,又可以像肿瘤细胞一样无限增殖。但是在二者融合过程中,除了B细胞与瘤细胞能融合外,B细胞自身、瘤细胞自身都能发生融合,此外还有多个B细胞与多个瘤细胞融合的情况(这种细胞能稳定存活的机率极小)以及未融合的细胞。B细胞或者B细胞自身融合后,仍然不能无限增殖,在增殖数代后就会死亡,因此主要问题转移到如何消除瘤细胞以及瘤细胞自身融合形成的细胞,整个单抗制备中,正是这一步让我们感受到那些科学家们的工作是多么炫丽。为了更好地理解他们是如何处理这个问题的,我们先来回顾一下生物化学中讲到的核酸的合成路线(以下图片截自王镜岩主编的生物化学教材)。
先来看嘌呤核苷酸的合成 生物体内先合成次黄嘌呤核苷酸,然后由次黄嘌呤核苷酸合成其它嘌呤核苷酸。次黄嘌呤核苷酸的合成路径如下:
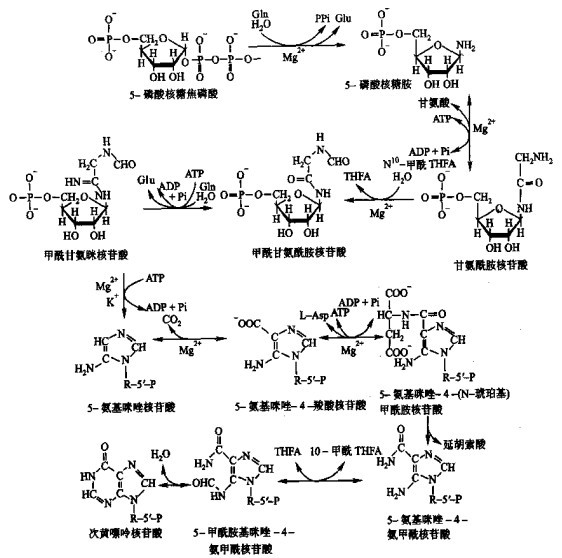
图1 次黄嘌呤核苷酸的合成
次黄嘌呤核苷酸合成后,经氨基化即可形成腺嘌呤核苷酸:
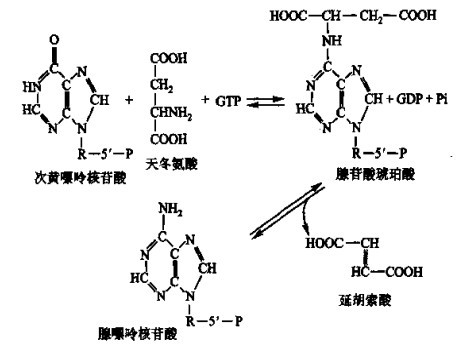
图2 次黄嘌呤核苷酸转化为腺嘌呤核苷酸
次黄嘌呤核苷酸合成后,经氧化可形成黄嘌呤核苷酸,黄嘌呤核苷酸经氨基化即可形成鸟嘌呤核苷酸:
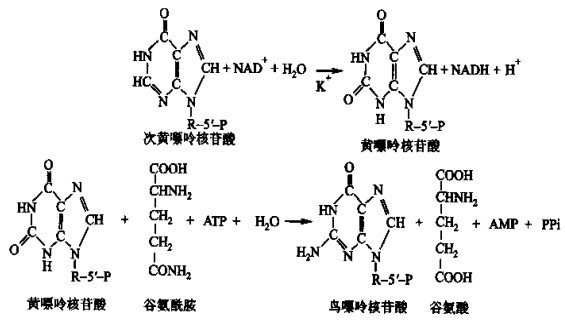
图3 次黄嘌呤核苷酸转化为鸟嘌呤核苷酸
再来看嘧啶核苷酸的合成 生物体内先合成尿嘧啶核苷酸,然后由尿嘧啶核苷酸转化为其它嘧啶核苷酸。尿嘧啶核苷酸的合成过程如下:
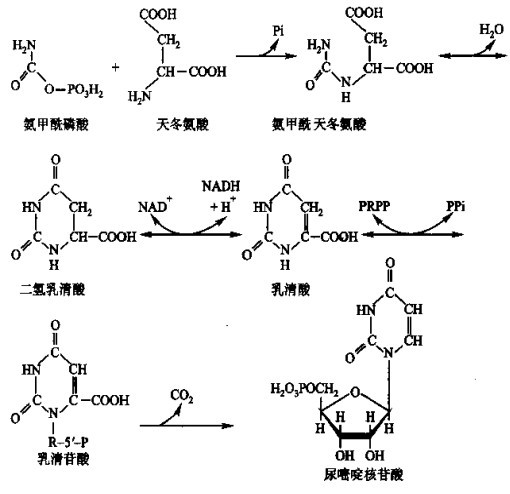
图4 尿嘧啶核苷酸的合成
尿嘧啶核苷酸经磷酸化形成尿嘧啶核苷三磷酸(UTP),UTP经氨基化就形成胞嘧啶核苷三磷酸(CTP):
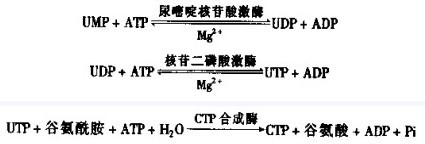
图5 胞嘧啶核苷三磷酸的合成
尿嘧啶核苷酸经核糖核酸还原酶还原后形成尿嘧啶脱氧核糖核苷酸。尿嘧啶脱氧核糖核苷酸经甲基化后形成胸腺嘧啶核苷酸:
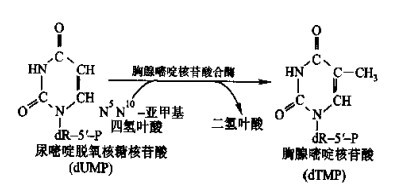
图6 胸腺嘧啶核苷酸的合成
脱氧核糖核核苷酸都可以由相应的核糖核苷酸经核糖核酸还原酶还原得到。
以上为核酸合成的主要途径。除此途径外,生物体内还有一条旁路合成途径(补救合成途径):

图7 核酸的旁路合成途径(补救合成途径)
关于核酸的合成暂时先讲到这里,接着回到前面的问题上来:如何消除未融合的瘤细胞以及瘤细胞自身融合得到的细胞。
科学先驱们是这样解决的:先用嘌呤类似物(如8-氮杂鸟嘌呤)对瘤细胞进行筛选,嘌呤结构类似物可以参与核酸的旁路合成(图7),但是合成出来的核酸不像正常的核酸那样具有完整的功能,因此这种细胞就会死去,于是存活下来的细胞都不能进行旁路合成核酸,其本质是旁路合成的次黄嘌呤鸟嘌呤磷酸核糖转移酶(HGPRT)突变,这种细胞被称为HGRPRT缺陷型(HGPRT-)。有了这种突变型的细胞,问题就比较容易解决了:由于B细胞和瘤细胞(HGPRT-)融合后既能进行主要途径合成核酸,也能进行旁路合成,而瘤细胞则只能进行主要途径合成核酸,所以只需要阻断核酸的主要途径,那么未融合的瘤细胞和瘤细胞自身融合的细胞将会死去,而只有B细胞和瘤细胞的融合细胞能利用旁路合成而得以存活。
那么,如何阻断核酸合成的主要途径呢?请回头看看嘌呤合成过程(图1的)第三个反应:甘氨酰胺核苷酸被甲基化为甲酰甘氨酰胺核苷酸,这一过程的甲基由N10-甲酰四氢叶酸完成,甲基化完成后它变为四氢叶酸,在酶的催化下,四氢叶酸可被甲酸经ATP活化而甲基化,重新回到N10-甲酰四氢叶酸。另外,胸腺嘧啶核苷酸的合成中(图6)也有类似的甲基化,此过程中,N5N10-亚甲基四氢叶酸转化为二氢叶酸。二氢叶酸经以下反应重新转化为N5N10-亚甲基四氢叶酸:

图8 N5N10-亚甲基四氢叶酸的重生
叶酸的结构类似物,如氨基蝶呤、氨甲蝶呤可以和二氢叶酸还原酶结合,从而阻止了二氢叶酸向N5N10-亚甲基四氢叶酸的转化,因此可以抑制胸腺嘧啶核苷酸的生成。另一方面,由嘌呤合成过程中产生的四氢叶酸可以产生图8中的第二个反应,所以四氢叶酸被逐渐消耗,也不能重生N10-甲酰四氢叶酸,于是嘌呤的合成路径也中断。这样就达到了阻止核酸主要途径合成的目的。因此,只需要在培养基中添加氨基蝶呤(A)(或氨甲蝶呤),同时加入次黄嘌呤(H)和胸腺嘧啶核苷酸(T)就可以选择出B细胞和瘤细胞的杂交瘤了,这也就是大家所知道的HAT选择培养基。
整个筛选原理就是这样的(示HAT作用,点击图片可查看高清大图):
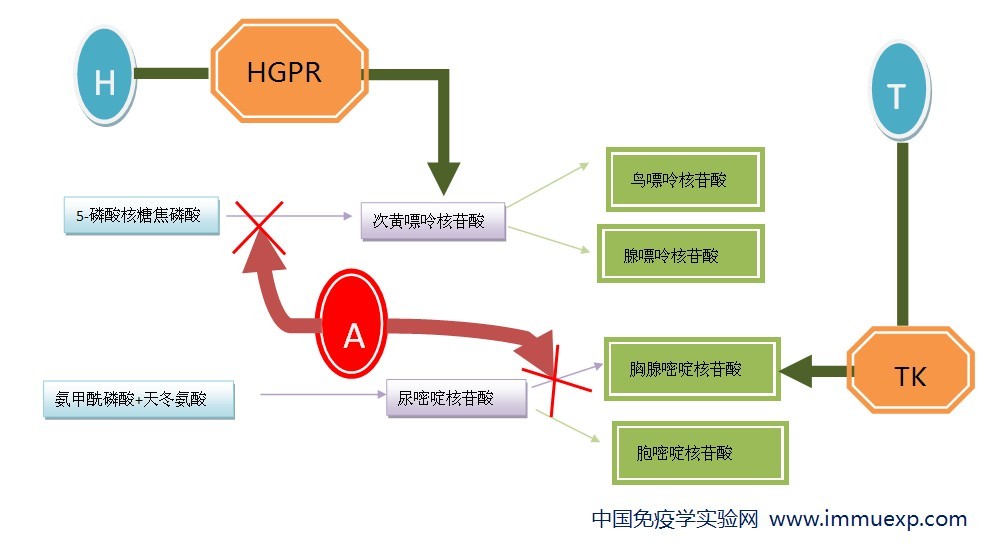
好了,到这里,筛选机制就讲完了,你可能会问:核酸的旁路合成中有那么多的酶参与反应,为什么只选择HGRPT酶缺陷作为筛选方法呢?原因是HGPRT酶的基因存在于X染色体上,而X染色体只有一条有活性,所以筛选出HGPRT酶缺陷比其它酶的缺陷来得更加容易了。(又一次对那些先驱表示膜拜!)当然,后来也有用胸苷激酶(TK)缺陷型作为筛选依据的。
除了小鼠单抗外,还有大鼠单抗、兔单抗,其制备原理和小鼠单抗类似,有兴趣的可以自己查阅文献或在本站论坛讨论。
概述部份就写这么多了,其实主要是瞻仰一下前辈们的光辉,和大家一起回顾一下单抗历程,其它每一步操作中涉及的原理将在每一部分独立讨论。
<----返回目录--->
关于这一部分如果还有什么问题可以在本站论坛上发帖(需要文献的也可以论坛提出,站长将陆续上传资料),也可以加入中国免疫学实验网QQ群和大家及时联系: 42654080。中国免疫学实验网感谢您的光临!
